Введите слово или словосочетание на любом языке 👆
Язык:
Перевод и анализ слов искусственным интеллектом ChatGPT
На этой странице Вы можете получить подробный анализ слова или словосочетания, произведенный с помощью лучшей на сегодняшний день технологии искусственного интеллекта:
- как употребляется слово
- частота употребления
- используется оно чаще в устной или письменной речи
- варианты перевода слова
- примеры употребления (несколько фраз с переводом)
- этимология
Что (кто) такое ПЕЧЕНЬ - В ПОРТАЛЬНАЯ ТРИАДА И АЦИНУС - определение
СТРУКТУРНАЯ ЕДИНИЦА ЛЁГКИХ
Лёгочный ацинус; Ацинус лёгкого
. 1, терминальная бронхиола; 2, респираторная бронхиола; 3, альвеолярный ход; 4, преддверие; 5, альвеолярный мешок; 6, альвеола, межальвеолярная перегородка: m, гладкая мускулатура; a, ветвь лёгочной артерии; v, ветвь лёгочной вены; s, перегородка между вторичными дольками.")
Найдено результатов: 16508
Агия-Триада

![1500]]—[[1400-е годы до н. э.]]. [[Археологический музей Ираклиона]]](https://commons.wikimedia.org/wiki/Special:FilePath/Harvester Rhyton.jpg?width=200 "1500]]—[[1400-е годы до н. э.]]. [[Археологический музей Ираклиона]]")
Айа-Триада; Агия Триада; Агиа Триада
Айия-Триас (Hagía Triás), древнее поселение на острове Крит, один из центров эгейской культуры (См. Эгейская культура). Названо по современной деревне А.-Т. Древнейшие остатки поселения относятся к неолиту, расцвет - к 1-й половине 2-го тыс. до н. э. Дома в А.-Т. группировались вокруг дворца, северо-западная часть которого хорошо сохранилась. Обнаружены залы, кладовые, помещение архива (судя по глиняным табличкам с письменами). Найдено много произведений искусства и ремесла. Упадок А.-Т. связан с нападением ахейцев на о. Крит (середина 15 в. до н. э.).
Лит.: Пендлбери Д., Археология Крита, пер. с англ., М., 1950.
Т. Д. Златковская.
Ацинус (лёгкое)
Ацинус ( «виноградная гроздь»Структура ацинуса по форме напоминает виноградную гроздь.) — структурно-функциональная единица лёгких.
печень

![[[Афлатоксин B1]] — чрезвычайно токсичен и сильнейший гепатоканцероген.](https://commons.wikimedia.org/wiki/Special:FilePath/(–)-Aflatoxin B1 Structural Formulae V.1.svg?width=200 "[[Афлатоксин B1]] — чрезвычайно токсичен и сильнейший гепатоканцероген.")


![крысы]]](https://commons.wikimedia.org/wiki/Special:FilePath/Perfused Rat Liver.JPG?width=200 "крысы]]")
НЕПАРНЫЙ ВНУТРЕННИЙ ОРГАН ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Печень человека
ж.
Самая крупная железа у животных и человека, вырабатывающая желчь.
Самая крупная железа у животных и человека, вырабатывающая желчь.
ПЕЧЕНЬ
НЕПАРНЫЙ ВНУТРЕННИЙ ОРГАН ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Печень человека
крупная железа у животных и человека, вырабатывающая желчь, участвующая в процессах пищеварения, кровообращение, обмена веществ.
ПЕЧЕНЬ
НЕПАРНЫЙ ВНУТРЕННИЙ ОРГАН ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Печень человека
самая большая железа в теле позвоночных. У человека она составляет около 2,5% от массы тела, в среднем 1,5 кг у взрослых мужчин и 1,2 кг у женщин. Печень расположена в правой верхней части брюшной полости; она прикрепляется связками к диафрагме, брюшной стенке, желудку и кишечнику и покрыта тонкой фиброзной оболочкой - глиссоновой капсулой. Печень - мягкий, но плотный орган красно-коричневого цвета и состоит обычно из четырех долей: большой правой доли, меньшей левой и гораздо меньших хвостатой и квадратной долей, образующих заднюю нижнюю поверхность печени.
См. также:
Печень
НЕПАРНЫЙ ВНУТРЕННИЙ ОРГАН ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Печень человека
крупная железа животного организма, участвующая в процессах пищеварения, обмена веществ, кровообращения и осуществляющая специфические защитные и обезвреживающие, ферментативные и выделительные функции, направленные на поддержание постоянства внутренней среды организма.
Сравнительная морфология. У беспозвоночных П. - пищеварительная железа, представляющая собой вырост средней кишки, принимает участие в переваривании и всасывании пищи; в ней откладываются запасные питательные вещества, жиры и углеводы. У многих беспозвоночных П. часто называется печёночно-поджелудочной железой (hepato-pancreas). У большинства моллюсков П. массивная, дольчатая, обычно парная; открывается в желудок 1-2 или многими протоками; клетки П. моллюсков способны к Фагоцитозу. Среди членистоногих П. имеют ракообразные, мечехвосты и большинство паукообразных. П. ракообразных - мешковидные выросты переднего отдела средней кишки; вырабатывает фермент, расщепляющий клетчатку. У паукообразных П. представлена парными выростами брюшного отдела средней кишки. Среди иглокожих большие печёночные выросты желудка имеются только у морских лилий и звёзд.
У хордовых животных и у человека П.- орган, в котором вырабатывается секрет, участвующий в пищеварении (жёлчь), и осуществляются важнейшие процессы, связанные с обменом веществ в организме. П. оболочников - обычно древовидно ветвящаяся железа; образуется как вырост желудка, тесно прилегает к его стенке и открывается в него одним протоком. У ланцетника П. представлена мешковидным печёночным выростом кишечника.
У позвоночных П. закладывается как брюшной вырост средней кишки, который, древовидно разветвляясь, приобретает строение трубчатой железы. Просветы конечных трубочек П. образуют жёлчные капилляры, по которым жёлчь стекает в более крупные печёночные протоки; отдельные печёночные протоки обычно объединяются в общий Жёлчный проток, открывающийся в двенадцатиперстную кишку (См. Двенадцатиперстная кишка). Обычно из части жёлчного протока образуется Жёлчный пузырь. Трубчатое строение П. сохраняется в течение всей жизни только у некоторых круглоротых (миксин). У миног, а также у рыб и земноводных трубчатое строение П. частично нарушается, так как между трубочками П. возникают поперечные перекладины - анастомозы и между ними врастает соединительная ткань с кровеносными сосудами и нервами. У пресмыкающихся, птиц и млекопитающих обильные анастомозы преобразуют трубчатую железу в сетчатую. У миног и у некоторых рыб П.- нерасчленённый орган, однако у большинства животных в ней имеются правая и левая лопасти (жёлчный пузырь всегда связан с правой лопастью). У некоторых животных (особенно среди млекопитающих) обе лопасти могут быть дольчатыми. У хищников П. относительно крупнее, чем у травоядных. У рыб и земноводных П.-крупнее, чем у пресмыкающихся, птиц и млекопитающих. Форма П. зависит от формы тела животного. У некоторых земноводных, рыб и млекопитающих П. тесно связана с поджелудочной железой (См. Поджелудочная железа), протоки которой открываются в жёлчный проток.
У человека П.- самая крупная пищеварительная железа. Развивается на 3-й неделе внутриутробной жизни плода из выроста эпителия слизистой оболочки двенадцатиперстной кишки. У взрослого человека П. весит 1,5-2 кг (около 1/50 массы тела), имеет плотную консистенцию, но меняет форму и размеры как от воздействия со стороны окружающих органов, так и от функциональных нагрузок. Сверху к П. прилежит диафрагма (см. Грудобрюшная преграда), над которой расположены сердце и лёгкие; снизу - желудок, двенадцатиперстная кишка, часть поперечной ободочной кишки, правая почка с надпочечником; сзади - пищевод и позвоночный столб; спереди - передняя стенка брюшной полости. На П. различают передневерхнюю, или диафрагмальную, и висцеральную (нижнюю) поверхности и края: передний - острый, задний - тупой. Передневерхняя выпуклая поверхность П. делится серповидной связкой на 2 доли - правую, большую, и левую, меньшую (рис. 1). Нижняя поверхность П. несколько вогнута. На ней различают правую и левую продольные борозды и поперечную (называют воротами П.), которые делят П. на 4 доли: собственно правую, левую, хвостатую, квадратную (рис. 2). В правой продольной борозде спереди лежит жёлчный пузырь, сзади - нижняя полая вена (См. Полые вены); в левой - спереди круглая связка П. (заросшая пупочная вена), сзади - венозная связка (заросшее соединение пупочной вены с нижней полой веной). В ворота П. входят воротная вена, печёночная артерия, нервы; выходят - лимфатические сосуды и печёночный проток, который, соединяясь с протоком жёлчного пузыря, образует жёлчевыносящий проток, впадающий в двенадцатиперстную кишку. П. покрыта серозной оболочкой (брюшиной (См. Брюшина)), за исключением участка, где она прилежит к диафрагмой срастается с ней. Серозная оболочка, переходя с П. на соседние органы, образует связки: серповидную, правую и левую венечные, соединяющие П. с диафрагмой, а также идущие от ворот П. печёночно-желудочную и др. Связки играют роль в удержании П. на месте. Однако большее значение в фиксации П. имеют: внутрибрюшное давление, обусловливающее более или менее плотное прилегание органов брюшной полости друг к другу и их взаимную опору; нижняя полая вена, плотно врастающая своими притоками (печёночными венами) в вещество П.; сила сцепления между серозными оболочками П. и диафрагмы в местах их соприкосновения; соединительная ткань, связывающая П. с диафрагмой в местах, не покрытых брюшиной. П. расположена основной массой в правом подреберье и через надчревную область простирается в левое подреберье. Нижняя граница П. в норме справа не выходит у взрослого человека из-под края правой рёберной дуги. У новорождённых П. занимает весь верхний отдел брюшной полости и левой долей касается селезёнки; нижний край П. нередко доходит до пупка, у грудного ребёнка он выступает на 2-3 см из-под рёберного края и только к 4 годам скрывается за ним.
П. - сложная трубчатая железа. Под её серозной оболочкой располагается соединительнотканная (глиссонова) капсула, содержащая эластические волокна; в воротах П. капсула утолщается и вместе с кровеносными сосудами проникает внутрь П., разделяя её на призматические печёночные дольки размером 0,5-2,0 мм (см. илл.). В середине каждой дольки проходит центральная вена, от которой по радиусам в виде перекладин (балок), называется печёночными пластинками, располагаются печёночные клетки. Печёночные пластинки в совокупности составляют железистую паренхиму П. Дольки П. состоят из тонких, но широких пластинок, анастомозирующих между собой и состоящих из одного слоя печёночных клеток. Между ними располагаются жёлчные капилляры, которые, сливаясь, образуют внутри- и междольковые жёлчные протоки, составляющие печёночный проток.
Кровоснабжение П. осуществляется по печёночной артерии, приносящей артериальную кровь, богатую кислородом, и воротной вене (см. Воротные системы). Кровь, поступающая в П. по воротной вене от желудка, селезёнки, кишечника, поджелудочной железы и др. органов брюшной полости, содержит некоторые продукты переваривания белков, углеводов и частично жиров, различные химические вещества, обеспечивающие физиологические функции П. Конечные ветви печёночной артерии и воротной вены внутри долек переходят в синусоиды, скорость кровотока в которых сравнительно низка. Здесь происходит обмен между кровью и клетками П., после чего кровь поступает в центральные вены, которые соединяются и в виде 3-4 печёночных вен впадают в нижнюю полую вену. Разветвленная капиллярная сеть, поверхность которой достигает 400 м; обеспечивает прохождение через П. около 2 тыс. л крови в сутки, причём 80\% её поступает по системе воротной вены, а 20\% - через печёночную артерию. Внутридольковые капилляры (синусоиды) в П., в отличие от обычных капилляров, снабжены, кроме эндотелия, ретикулярными клетками звездчатой формы (см. Купфера клетки), которые относятся к ретикуло-эндотелиальной системе (См. Ретикуло-эндотелиальная система) и обладают выраженной способностью к фагоцитозу. Иннервируется П. блуждающими нервами и ветвями солнечного сплетения.
Физиология и биохимия. Физиологическое значение П. определяется тем, что всасываемые из кишечника в кровь вещества проходят через П. и подвергаются в ней химическим изменениям. П. участвует в поддержании динамического равновесия многих веществ плазмы крови (сахара, холестерина, белков крови, железа, ретинола, или витамина А, воды). Через П. протекает в 1 мин около 1,5 л крови, в ней освобождается 1/7 часть всей энергии организма. Температура оттекающей от неё крови во время пищеварения увеличивается на 1-2 °С. П. инактивирует многие гормоны: тироксин, эстрогены, гонадотропные гормоны, стероиды коры надпочечников, серотонин и др. Некоторые вещества, пройдя через П., приобретают большую токсичность, например алкалоид колхицин превращается в более ядовитое вещество - оксиколхицин; сульфаниламиды после ацетилирования в П. становятся менее растворимыми, вследствие чего осаждаются в мочевых путях. П. обладает жёлчеобразовательной функцией. Жёлчь, синтезируемая в клетках П. из веществ, поступающих с кровью, имеет важное значение для процессов пищеварения (См. Пищеварение) и жирового обмена (См. Жировой обмен) в организме. Не менее важная функция П.- депонирование крови (см. Депо крови). Сосуды П. могут вмещать 20\% всей крови, поэтому задержка крови в нормальной здоровой П. не представляет собой венозного застоя. От её функционального состояния зависит деятельность др. депонирующих кровь органов (селезёнки, кишечника). Вся кровь, вышедшая из селезёнки и кишечника, обязательно проходит через П. Здесь удаляется избыток воды из крови, который идёт на создание лимфы и жёлчи. В П. образуется от 1/3 до 1/2 всей лимфы с большим содержанием белка (6\%).
В состав П. входят: вода (70-75\%), простые и сложные белки (12-24\%) и продукты их распада, липиды (2-6\%), углеводы (2-8\%) и продукты их расщепления, коферменты, витамины, гормоны, разнообразные низкомолекулярные органические вещества и минеральные катионы и анионы. П. выполняет весьма важные функции. В ней проходят процессы биосинтеза важнейших для организма соединений - нуклеиновых кислот (См. Нуклеиновые кислоты) (ДНК и РНК), различных ди- и мононуклеотидов, пуриновых и пиримидиновых оснований. В то же время ферменты, содержащиеся в П., вызывают расщепление нуклеиновых кислот и нуклеотидов, дезаминирование и окисление свободных пуриновых оснований. П. участвует в той или иной степени в обмене белков, углеводов, липидов, витаминов, минеральных веществ и воды. Продукты расщепления всех питательных веществ образуют в П. основной "метаболический фонд", из которого организм черпает по мере надобности необходимые для него вещества.
Белковый обмен. Из 80-100 г белка, расщепляемого и вновь синтезируемого в организме человека за сутки, примерно половина приходится на П. Белки в П. обновляются за 7 суток, а в др. органах - за 17 и более. Это свидетельствует об интенсивности белкового обмена в П. В ней происходит синтез белков, начиная с активации аминокислот в гиалоплазме, образования соединений со специфическими для каждой аминокислоты транспортными РНК и кончая завершающей стадией синтеза - высвобождением длинных пептидных цепей готовых белков из места их синтеза в рибосомах. В П. образуются не только белки, характерные для неё самой, но и белки плазмы крови - альбумины, многие глобулины, а также фибриноген и др. факторы, участвующие в процессе свёртывания крови (См. Свёртывание крови). Под влиянием катептических протеаз и пептидаз (см. Катепсины.) в П. происходит расщепление белков и образование аминокислот, которые подвергаются в ней различным превращениям: дезаминированию (практически происходит только в П.), переаминированию, декарбоксилированию, приводящему к возникновению биогенных аминов; в результате переноса метильной группы от аденозилметионина обеспечивается образование холина, креатина, адреналина и др. метилированных соединений. Своеобразны и характерны пути превращения в П. отдельных аминокислот - триптофана, фенилаланина, гистидина, лизина и др. Из триптофана, в частности, синтезируются такие биологически активные вещества, как триптамин, окситриптофан и продукт его декарбоксилирования - Серотонин, хинолиновая кислота и оба продукта её декарбоксилирования - никотиновая и пиколиновая кислоты; из гистидина образуются формиминоглутаминовая и глутаминовая кислоты, а также Гистамин; из аргинина образуются орнитин и мочевина. Орнитин вступает в характерный цикл реакций, приводящих к синтезу конечного продукта обмена простых белков - мочевины (из углекислого газа и аммиака при участии ионов магния, АТФ и ряда аминокислот - см. Орнитиновый цикл). В П. происходят синтезы, нейтрализующие токсические продукты обмена веществ, такие, как фенолы, ароматические углеводороды и многие др. с образованием гиппуровой и фенацетуровой кислот (при использовании глицина), а также парных глюкуроновых эфиросерных кислот, меркаптокислот и др. соединений.
Углеводный обмен. П. поддерживает концентрацию сахара в крови на таком уровне, который обеспечивает непрерывное снабжение глюкозой (См. Глюкоза) всех тканей. Это достигается регуляцией соотношения между синтезом и распадом Гликогена, депонируемого в П. (см. Кори цикл). В среднем П. человека содержит 30-100 г гликогена. Этого количества достаточно, чтобы служить резервуаром для регуляции уровня сахара в крови. При всасывании сахара из кишечника содержание глюкозы в крови воротной вены может повышаться до 400 мг\%, а в периферической крови её содержится не более 200 мг\% Глюкоза превращается в П. в гликоген и депонируется, а также используется для получения энергии. Если после этого и др. синтезов ещё имеется избыток глюкозы, она превращается в жир. При голодании П. поддерживает постоянный уровень сахара в крови прежде всего расщеплением гликогена; если этого недостаточно - гликонеогенезом (превращением гликогенных аминокислот и глицерина в сахар). Инсулин, образующийся во внутрисекреторных отделах поджелудочной железы, проходя через П., также оказывает влияние на уровень сахара в крови и на образование и распад гликогена в П. Под влиянием фосфорилазы концевые глюкозные остатки гликогена отщепляются с образованием глюкозо-1-фосфата, участвующего в образовании уридиндифосфатглюкозы - транспортной формы глюкозных остатков и основного их источника при синтезе гликогена. Нарушение ферментативного превращения галактозо-1-фосфата в глюкозо-1-фосфат приводит к тяжёлым патологическим явлениям, связанным с наследственной болезнью - галактоземией. Обычный путь превращения глюкозо-1-фосфата (образование из него глюкозо-6-фосфата) имеет большое биологическое значение, так как это соединение играет центральную роль в превращениях углеводов и саморегуляции углеводного обмена (См. Углеводный обмен). В П. глюкозо-6-фосфат резко тормозит фосфоролитическое расщепление гликогена, активирует ферментативный транспорт глюкозы с уридинфосфоглюкозы на молекулу строящегося гликогена, является субстратом для окислительного превращения глюкозы по пентозофосфатному пути. При окислении глюкозо-6-фосфата образуется восстановленная форма Никотинамидадениндинуклеотидфосфата (НАДФ) - необходимого кофермента восстановительных синтезов жирных кислот и холестерина и превращения глюкозо-6-фосфата в фосфопентозы - обязательный компонент при образовании нуклеотидов и нуклеиновых кислот. Кроме того, глюкозо-6-фосфат - субстрат для дальнейших гликолитических превращений, приводящих через фруктозомоно- и дифосфаты к фосфотриозам и образованию пировиноградной и молочной кислот. Этот процесс обеспечивает организм соединениями, необходимыми для биосинтезов, и играет существенную роль в обмене энергии, так как образование каждой молекулы молочной кислоты равноценно синтезу одной богатой энергией фосфатной связи в молекуле АТФ. Наконец, расщепление глюкозо-6-фосфата фосфатазой обеспечивает поступление в кровь свободной глюкозы, доставляемой током крови во все органы и ткани.
Жировой обмен. П. в состоянии депонировать значительно больше липидов, чем гликогена (до 20-30\% сухой массы). Часть липидов, состоящая из фосфатидов и холестерина, довольно постоянна и составляет 10-15\%; содержание нейтрального жира колеблется. Депонирование жира - функция жировой ткани, а не П. В целом П. не играет в липидном отмене такой жизненно важной роли, как в углеводном и белковом обменах. Расщепление жирных кислот также не ограничивается П. В П. происходят деградация жира и окисление жирных кислот, а также представлены ферментные системы биосинтеза высокомолекулярных жирных кислот, нейтрального жира и сложных липидов; промежуточный продукт при этих синтезах - фосфатидная кислота. В П. синтезируется также холестерин. Образующиеся при деградации жира жирные кислоты окисляются с образованием ацетилкофермента A, вступающего при наличии конденсирующего фермента в реакцию со щавелевоуксусной кислотой и образующего таким образом лимонную кислоту - основной субстрат окислительных превращений в Трикарбоновых кислот цикле. В клетках П., как и в клетках др. органов, окислительного превращения, локализованные по преимуществу в митохондриях, сопряжены с образованием богатых энергией соединений (АТФ) и заканчиваются образованием CO2 и H2O. Синтез высокомолекулярных жирных кислот протекает вне митохондрий - в так называемом цитозоле и, следовательно, пространственно отделен от места их окисления. В так называемой микросомальной фракции П. сосредоточена 2-я (не митохондриальная) НАДФ-зависимая система окисления углеводородов, стероидов, холестерина. Эта система локализована в эндоплазматическом ретикулуме и связана с образованием продуктов гидроксилирования. П. имеет существенное значение в об мене пигментов: в ней разрушается Гемоглобин, образуется билирубин и превращается в растворимую форму в виде диглюкуронида билирубина. Пигментный обмен в П., тесно связанный с метаболизмом билирубина и порфиринов, в свою очередь, играет важную роль в обмене железа в организме.
В минеральном обмене и в сохранении постоянства кислотно-щелочного равновесия (См. Кислотно-щелочное равновесие) П. принимает непосредственное участие. Минеральные вещества в П. находятся как в свободном виде, так и входят в состав сложных органических соединений, например ферментов (Mg, Mn, Fe, Cu, Zn). Катионы выполняют также роль активаторов ферментов, например Na+, Ca2+, К+, Ni2-, Co2+, Cr3+ и др. В составе П. находятся железосодержащий белок ферритин и медьсодержащий белок гепатокупреин; эти вещества участвуют в процессе кроветворения (См. Кроветворение). П. также участвует в обмене витаминов. В ней содержатся витамины группы В и D, витамин C и растворимые в жирах витамины E и K. Из каротинов в П. образуется и депонируется витамин A, всасывание которого из кишечника происходит только в присутствии жёлчи. Аскорбиновая кислота способствует гликогенезу в П. Витамин К необходим для синтеза Протромбина в ней.
Функции П. (процессы обмена веществ, протекающие в ней, жёлчеобразование) регулируются нервными и гормональными механизмами. В гормональной регуляции участвуют адреналин, инсулин, глюкагон, кортикостероиды, гормоны, вырабатываемые в гипофизе, интестинальные гормоны, особенно секретин, холецистокинин, панкреозимин. Влияние многих гормональных факторов реализуется в П. при участии циклических мононуклеотидов: циклического аденозинмонофосфата (цАМФ) и гуанозинмонофосфата (цГМФ). Эти циклические мононуклеотиды образуются при расщеплении циклазой (ферментом, фиксированным главным образом в плазматической мембране) нуклеозидтрифосфатов АТФ и ГТф. Циклические мононуклеотиды выполняют функции регуляторов активности многих ферментов в результате активации протеинкиназ, обеспечивающих процесс переноса фосфатного остатка с АТФ на белки-ферменты. Фосфорилирование ферментов меняет их активность, повышая её у одних (фосфорилазы, липазы) и подавляя у других (гликоген-синтетазы, пируватдекарбоксилазы).
В свою очередь, биохимические процессы, протекающие в П., через тканевые интероцепторы оказывают влияние на функциональное состояние центральной нервной системы. Разнообразие и взаимозависимость факторов, влияющих на состояние клеток П., обусловливают как интенсивность, так и направленность протекающих в ней процессов обмена веществ.
При заболеваниях П. у человека и животных поражается преимущественно её паренхима (клетки) или межуточная ткань. Острые Гепатиты составляют значительную часть всех болезней П. и могут быть причиной развития хронических её поражений. Среди гепатитов инфекционной природы различают первичные (см. Гепатит вирусный) и вторичные (например, при бруцеллёзе (См. Бруцеллез), лептоспирозе (См. Лептоспирозы), Сифилисе и др.). Токсико-аллергические гепатиты развиваются при воздействии на организм химических, в том числе лекарственных веществ (см. Аллергия). Исходом болезни может быть Цирроз печени. В результате нарушения питания и обмена веществ (при Алкоголизме, витаминной недостаточности (См. Витаминная недостаточность), патологическом голодании (См. Голодание), диабете сахарном (См. Диабет сахарный), гепато-церебральной дистрофии (См. Гепато-церебральная дистрофия), ожирении (См. Ожирение) и т.д.), а также при нарушениях кровообращения, особенно при повышении давления в нижней полой и печёночной венах, в П. развиваются дистрофические изменения. П. человека и животных может быть местом обитания многих паразитов: простейших, гельминтов, реже членистоногих. Так, из кишечника в жёлчные пути могут проникать лямблии, лейшмании (см. Лямблиоз, Лейшманиоз); в П. человека происходит внеэритроцитарное развитие возбудителя малярии (См. Малярия). В протоках П. и желчном пузыре паразитируют различные трематоды, вызывающие Холангиты, Холециститы (см. Дикроцелиоз, Клонорхоз, Описторхоз, Шистосоматозы, Фасциолёз). П. может быть местом развития возбудителей Эхинококкоза. Из новообразований П. встречаются рак (См. Рака), Саркома и др.
Лит.: Догель В. А., Сравнительная анатомия беспозвоночных, ч. 1, Л., 1938; Шмальгаузен И. И., Основы сравнительной анатомии позвоночных животных, 4 изд., М., 1947; Павлов И. П., Лекции по физиологии, Полн. собр. соч., 2 изд., т. 5, М.- Л., 1952; Фишер А., Физиология и экспериментальная патология печени, пер. с англ., Будапешт, 1961; Рапопорт С. М., Медицинская биохимия, пер. с нем., М., 1966; Введение в клиническую биохимию (основы патобиохимии), Л., 1969: Бондарь З. А., Клиническая гепатология, М., 1970; Блюгер А. Ф., Райцис А. Б., Серотонин и печень, "Успехи гепатологии", в. 3, Рига, 1971; Збарский Б. И., Иванов И. И., Мардашев С. Р., Биологическая химия, 5 изд., Л., 1972.
С. Е. Северин, А. Н. Дружинин, А. А. Гладышева.
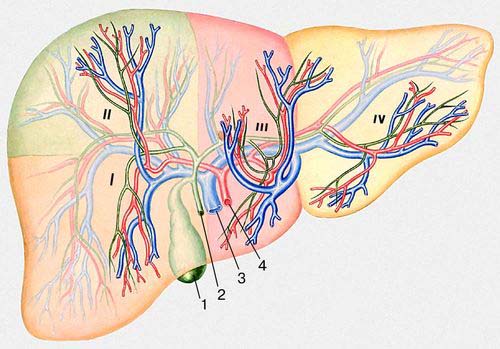
Основные сегменты печени человека (вид спереди): I - передний, II - задний, III - медиальный, IV - латеральный; 1 - желчный пузырь; 2 - жёлчевыводящий проток; 3 - воротная вена; 4 - печёночная артерия.
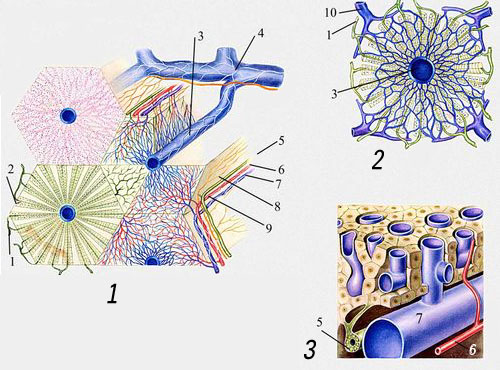
1-3. Схемы строения печёночной дольки: 1 - жёлчные ходы; 2 - жёлчные капилляры; 3 - центральная вена; 4 - поддольковая (собирательная) вена; 5 - междольковый проход; 6 - междольковая артерия; 7 - междольковая вена; 8 - междольковые лимфатические капилляры; 9 - нервное сплетение вокруг сосуда; 10 - приток междольковых вен.
Рис. 1. Печень человека (вид спереди и сверху): 1 - диафрагма; 2 - венечная связка печени; 3 - левая треугольная связка; 4 - левая доля; 5 - серповидная связка; 6 - круглая связка; 7 - передний край; 8 - жёлчный пузырь; 9 - правая доля; 10 - правая треугольная связка.
Рис. 2. Печень человека (вид снизу): 1 - левая доля; 2 - хвостатая доля; 3 - нижняя полая вена; 4 - задняя поверхность; 5 - почечное вдавление; 6 - место перехода брюшины на печень; 7 - правая доля; 8 - вдавленне ободочной кишки; 9 - жёлчный пузырь; 10 - квадратная доля; 11 - круглая связка; 12 - пузырный проток; 13 - жёлчевыносящий проток; 14 - печёночный проток; 15 - воротная вена; 16 - печёночная артерия; 17 - венозная связка; 18 - желудочное вдавление.
печень
НЕПАРНЫЙ ВНУТРЕННИЙ ОРГАН ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Печень человека
ПЕЧЕНЬ
НЕПАРНЫЙ ВНУТРЕННИЙ ОРГАН ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Печень человека
крупная железа у животных и человека; участвует в процессах пищеварения, обмена веществ, кровообращения; обеспечивает постоянство внутренней среды организма. У позвоночных животных и человека клетки печени синтезируют желчь. В печени происходит синтез и расщепление белков, липидов, углеводов (регулирует уровень сахара в крови), витаминов (образуется и накапливается витамин А) и других веществ. Из "обменного фонда" печени организм получает многие необходимые вещества; в ней освобождается 1/7 часть всей его энергии. Через печень протекает в 1 мин ок. 1,5 л крови; в сосудах печени может находиться до 20% объема всей циркулирующей крови.
Печень
НЕПАРНЫЙ ВНУТРЕННИЙ ОРГАН ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Печень человека
Пе́чень (, ) — жизненно важная железа внешней секреции позвоночных животных, в том числе и человека, находящаяся в брюшной полости (полости живота) под диафрагмой и выполняющая большое количество различных физиологических функций. Печень является самой крупной железой позвоночных.
ПЕЧЕНЬ - В. ПОРТАЛЬНАЯ ТРИАДА И АЦИНУС
К статье ПЕЧЕНЬ
Ветви воротной вены, печеночной артерии и желчного протока расположены рядом, у наружной границы дольки и составляют портальную триаду. На периферии каждой дольки находится несколько таких портальных триад.
Функциональной единицей печени считается ацинус. Это - часть ткани, которая окружает портальную триаду и включает лимфатические сосуды, нервные волокна и прилегающие секторы двух или более долек. Один ацинус содержит около 20 печеночных клеток, расположенных между портальной триадой и центральной веной каждой дольки. В двумерном изображении простой ацинус выглядит как группа сосудов, окруженная прилегающими участками долек, а в трехмерном - похож на ягоду (acinus - лат. ягода), висящую на стебельке из кровеносных и желчных сосудов. Ацинус, микрососудистый каркас которого состоит из перечисленных выше кровеносных и лимфатических сосудов, синусоидов и нервов, является микроциркуляторной единицей печени.
Клетки печени (гепатоциты) имеют форму многогранников, но основных функциональных поверхностей у них три: синусоидальная, обращенная в синусоидальный канал; канальцевая - участвующая в образовании стенки желчного капилляра (собственной стенки он не имеет); и межклеточная - непосредственно граничащая с соседними печеночными клетками.
Википедия
Ацинус (лёгкое)

Ацинус (лат. acinus «виноградная гроздь») — структурно-функциональная единица лёгких. Состоит из ветвей терминальной (концевой) бронхиолы — респираторных бронхиол 19-23-го порядков и альвеолярных ходов, разветвляющихся на альвеолярные мешочки.

